
Il sonno REM, la “sentinella” del nostro riposo
Il sonno è uno stato comportamentale reversibile, caratterizzato da un’elevata soglia di attivazione e da una transitoria disconnessione sensoriale dall’ambiente esterno. Il sonno è un comportamento altamente conservato durante l‘evoluzione, presente anche in invertebrati privi di un cervello propriamente detto, a testimonianza dell’importanza cruciale delle sue funzioni (Aime, 2022).
La ricerca scientifica ha evidenziato un ruolo del sonno nei processi di sviluppo cerebrale e di detossificazione cerebrale (rimozione delle sostanze di scarto prodotti dal metabolismo cerebrale), oltre che nei meccanismi di plasticità sinaptica, alla base della nostra flessibilità comportamentale e della nostra capacità di apprendere nuove informazioni (Weiss & Donlea, 2022).
Tali benefici sono però ottenuti al prezzo di un ridotto monitoraggio dell’ambiente durante il sonno, che comporta il pericolo di aggressioni o di esposizione a pericoli ambientali.
In particolare, si è visto che la predazione è un importante determinante dell’architettura del sonno, e le specie animali che vivono in ambienti ad alto rischio di predazione (come topi e conigli) dormono meno di specie predatorie come i carnivori (Allison & Cicchetti, 1976).
Gli animali affrontano costantemente minacce poste da predatori, fattori ambientali o conspecifici aggressivi, e i loro cervelli hanno sviluppato sistemi di difesa attivati dagli input sensoriali che rilevano le minacce (LeDoux, 2012). I segnali olfattivi o visivi del predatore comunemente innescano una risposta che comprende un aumento dell’arousal e comparsa di un comportamento difensivo.
Come detto, durante il sonno la reattività comportamentale agli stimoli sensoriali risulta ridotta, così come la capacità di produrre risposte adeguate alle minacce ambientali. Negli anni ’60 Snyder ipotizzò che il sonno REM potesse fungere da “sentinella” durante il sonno, favorendo la periodica comparsa di brevi risvegli che consentono di mettere in atto una risposta immediata di “combattimento o fuga”; questi brevi risvegli garantirebbero quindi la massima protezione verso pericoli esterni, con un disturbo della continuità del sonno ridotto al minimo (Snyder, 1966).
In accordo con questa ipotesi, tutti gli animali, compresi gli esseri umani, sono più vigili dopo un risveglio da sonno REM rispetto ad un risveglio da sonno non REM, che è seguito da un lungo periodo di inerzia del sonno, uno stato fisiologico di compromissione delle prestazioni cognitive e sensoriali-motorie che compare immediatamente dopo il risveglio (Hilditch, 2019).
L’articolo pubblicato su “Neuron”
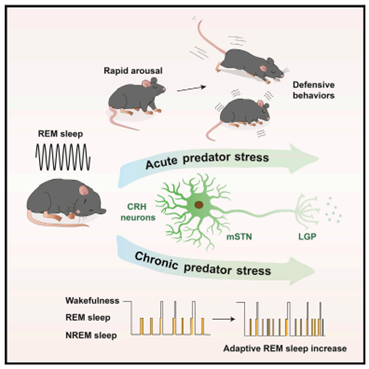
Un bell’articolo pubblicato quest’anno su Neuron da Tseng e collaboratori (Tseng et al, 2022) ha finalmente fornito una elegante dimostrazione sperimentale dell’ipotesi del sonno REM “sentinella” avanzata da Snyder. In questo lavoro, per prima cosa, gli Autori hanno dimostrato che gli animali (topi) esposti all’odore del predatore si svegliano con facilità molto maggiore dal sonno REM piuttosto che dal sonno non REM. Il risveglio è seguito da una rapida ed intensa risposta di arousal.
Quindi, hanno caratterizzato i circuiti neurali specifici in grado di indurre una risposta di arousal e comportamento difensivo. In particolare, hanno indagato il ruolo dei neuroni del nucleo subtalamico mediale (mSTN). mSTN fa parte dei circuiti dei gangli della base, che coordinano risposte emozionali, cognitive e motorie, e sono anche implicati nella regolazione del ciclo veglia-sonno (Vetrivelan et al, 2010). Questo nucleo contiene un numero elevato di neuroni che esprimono CRH (corticotropin-releasing hormone), un mediatore della risposta allo stress.
Gli Autori hanno dimostrato che durante il sonno REM l’esposizione a stimoli olfattivi di predatori attiva fortemente i neuroni mSTN-CRH, con un pattern di attivazione del tutto analogo a quello che si ha quando l’esposizione viene fatta durante la veglia. L’attivazione non si manifesta invece se l’esposizione avviene durante il sonno non REM. L’inibizione (optogenetica o chemogenetica) dei neuroni mSTN-CRH prolunga la latenza del risveglio da sonno REM e aumenta i tempi di reazione allo stimolo predatorio.
Al contrario, durante il sonno REM, la stimolazione optogenetica di questi neuroni promuove la veglia se sono presenti stimoli olfattivi predatori, ma consolida il sonno se questi sono assenti; ciò suggerisce che le proiezioni dei neuroni mSTN-CRH possano stabilizzare il sonno quando le condizioni ambientali sono sicure. Se l’esposizione a condizioni ambientali minacciose si protrae per un periodo prolungato, si ha una frammentazione del sonno ed un incremento del sonno REM. La frammentazione del sonno, con un sostanziale mantenimento della sua durata complessiva, consente da un lato di preservarne le funzioni, dall’altro di aumentare la probabilità dell’organismo di rilevare i pericoli ambientali e sfuggirli.
L’aumento della durata del sonno REM e del numero dei suoi episodi aumenta la capacità dell’animale di emergere rapidamente dal sonno, e garantisce livelli di vigilanza più elevati al risveglio. In questo modo il sonno REM svolge il suo ruolo di “sentinella” e consente un monitoraggio efficace dell’ambiente ostile.
Il sonno REM “sentinella”: conclusioni e prospettive future
In prospettiva, tutti gli studi che indagano i meccanismi cerebrali alla base della regolazione della veglia e del sonno nell’ambiente naturale, in condizioni di pericolo, come in questa ricerca, ma anche in circostanze particolari, come le migrazioni o l’ibernazione, sono cruciali per la comprensione degli adattamenti comportamentali a nicchie ambientali particolari, che indirizzano i processi di selezione naturale.
L’applicazione traslazionale di queste conoscenze agli esseri umani, e alla nostra organizzazione sociale potrà poi sicuramente migliorare il rilievo, la prevenzione e il trattamento dei disturbi del sonno, come anche di tutte le perturbazioni del sonno associate a condizioni neuropatologiche quali demenza, schizofrenia e depressione.
Giovanna Zoccoli, Commissione Divulgazione AIMS
Riferimenti Bibliografici
Aime M & Adamantidis AR (2022) Sleep to survive predators. Neurosci. Bull, 38:1114–1116
Allison T & Cicchetti DV (1976) Sleep in mammals: Ecological and constitutional correlates. Science 1976, 194: 732–734.
Hilditch CJ & McHill AW (2’19) Sleep inertia: Current insights. Nat Sci Sleep, 11:155–165.
LeDoux, J (2012). Rethinking the emotional brain. Neuron 73:653–676.
Snyder F (1966) Toward an evolutionary theory of dreaming. Am J Psychiatry, 123:121–142.
Tseng YT, Zhao BH, Chen SP et al. (2022) The subthalamic corticotropin-releasing hormone neurons mediate adaptive REM-sleep responses to threat. Neuron, 110:1223-1239.e8.
Vetrivelan R, Qiu MH, Chang C, & Lu J (2010). Role of basal ganglia in sleep-wake regulation: neural circuitry and clinical significance. Front. Neuroanat. 4:145.
Weiss JT & Donlea JM (2022) Roles for sleep in neural and behavioral plasticity: Reviewing variation in the consequences of sleep loss. Front Behav. Neurosci. 15:777799.